
Spatial Ecology and Diel Activity of European Wildcat (Felis silvestris) in a Protected Lowland Area in Northern Greece

 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
- (a)
- estimate the home range of two male and three female wildcats in Koronia and Volvi Lakes National Park (NPKV) in northern Greece,
- (b)
- describe the diel activity of both sexes,
- (c)
- identify potential environmental variables (e.g., weather, day length, moon phase) significantly affecting wildcat activity,
- (d)
- evaluate whether non-resting wildcat activity differs in open vs. forested areas, and
- (e)
- ascertain resting wildcat site fidelity across seasons.
2. Materials and Methods
2.1. Study Area
2.2. Trapping and Handling
2.3. GPS Collaring
2.4. Home Range Estimation
2.5. Activity Pattern Analysis
3. Results
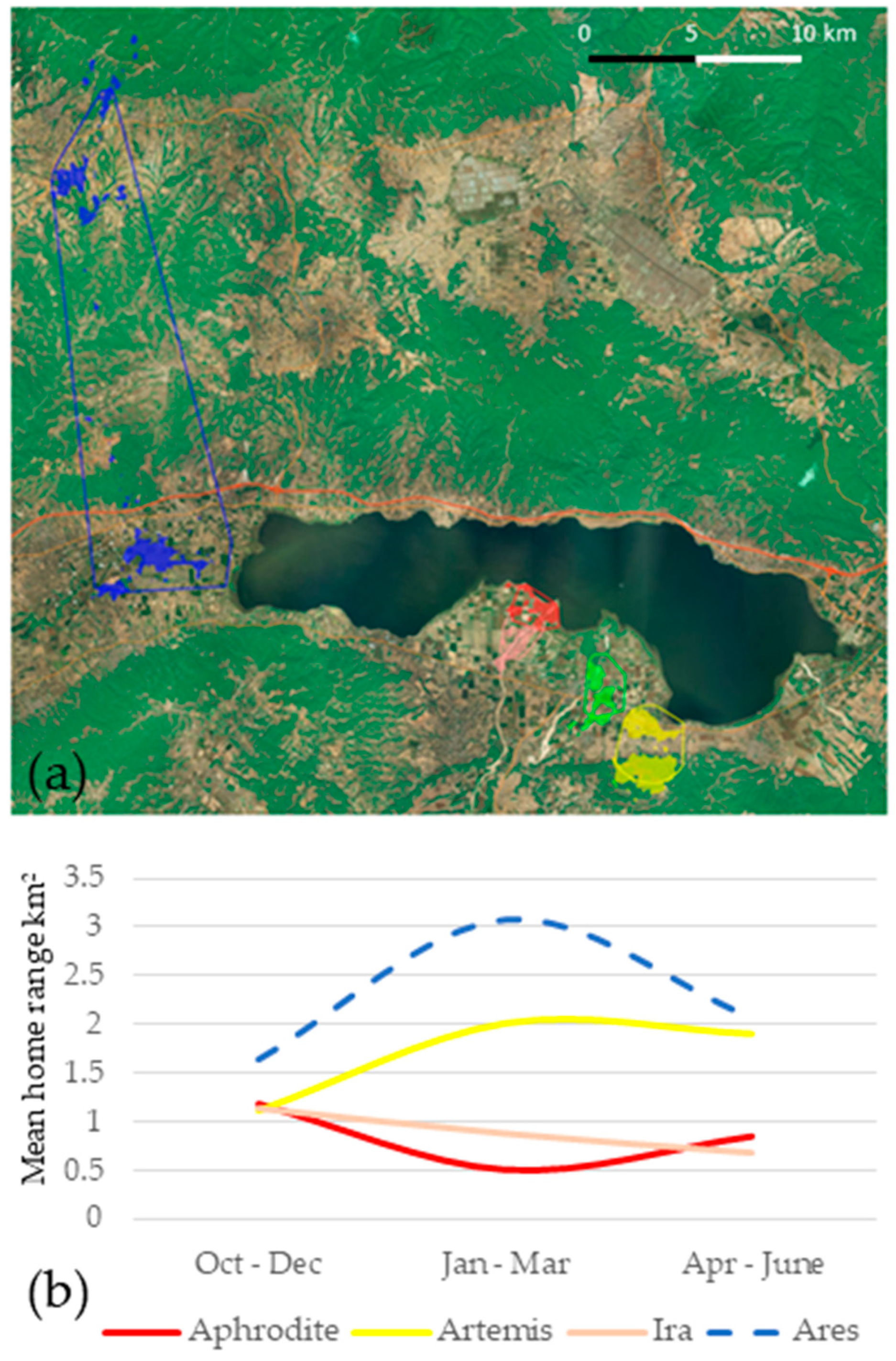
3.1. Home Range
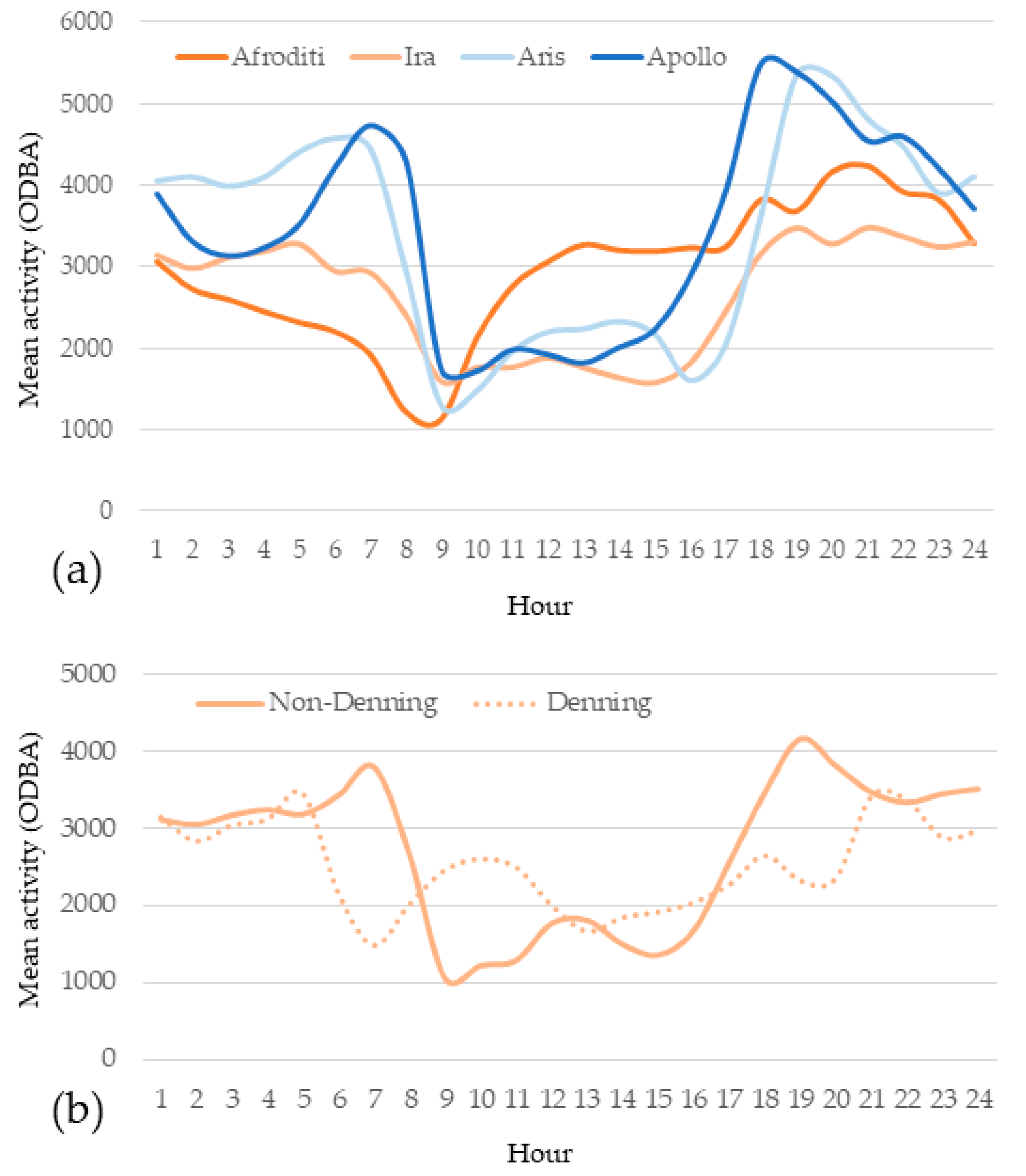
3.2. Activity Pattern
3.3. Resting Sites
3.4. Activity Differences in Open and Forested Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gerngross, P.; Ambarli, H.; Angelici, F.M.; Anile, S.; Campbell, R.; Ferreras, P.; Gil-Sanchez, J.M.; Götz, M.; Jerosch, S.; Mengulluoglu, D.; et al. Felis Silvestris. In IUCN Red List of Threatened Species; IUCN: Grand, Switzerland, 2021; submitted. [Google Scholar]
- Kitchener, A.C.; Breitenmoser-Würsten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A Revised Taxonomy of the Felidae: The Final Report of the Cat Classification Task Force of the IUCN/SSC Cat Specialist Group. Special Issue 11. Cat News, 2017; 80p. Available online: https://repository.si.edu/bitstream/handle/10088/32616/A_revised_Felidae_Taxonomy_CatNews.pdf(accessed on 20 August 2021).
- Yamaguchi, N.; Kitchener, A.; Driscoll, C.; Nussberger, B. Felis Silvestris. In IUCN Red List of Threatened Species; E.T60354712A50652361; IUCN: Grand, Switzerland, 2015. [Google Scholar] [CrossRef] [Green Version]
- Lozano, J.; Virgós, E.; Malo, A. Importance of Scrub–Pastureland Mosaics for Wild-Living Cats Occurrence in a Mediterranean Area: Implications for the Conservation of the Wildcat (Felis Silvestris). Biodivers. Conserv. 2003, 12, 921–935. [Google Scholar] [CrossRef]
- Anile, S.; Devillard, S.; Ragni, B.; Rovero, F.; Mattucci, F.; lo Valvo, M. Habitat Fragmentation and Anthropogenic Factors Affect Wildcat Felis Silvestris Silvestris Occupancy and Detectability on Mt Etna. Wildl. Biol. 2019, 2019, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gil-Sánchez, J.M.; Barea-Azcón, J.M.; Jaramillo, J.; Herrera-Sánchez, F.J.; Jiménez, J.; Virgós, E. Fragmentation and Low Density as Major Conservation Challenges for the Southernmost Populations of the European Wildcat. PLoS ONE 2020, 15, e0227708. [Google Scholar] [CrossRef]
- Klar, N.; Herrmann, M.; Kramer-Schadt, S. Effects and Mitigation of Road Impacts on Individual Movement Behavior of Wildcats. J. Wildl. Manag. 2009, 73, 631–638. [Google Scholar] [CrossRef]
- Lozano, J.; Malo, A.F. Conservation of the European wildcat (Felis silvestris) in mediterranean environments: A reassessment of current threats. In Mediterranean Ecosystems: Dynamics, Management and Conservation; Williams, G.S., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2012; pp. 1–31. [Google Scholar]
- Bastianelli, M.L.; Premier, J.; Herrmann, M.; Anile, S.; Monterroso, P.; Kuemmerle, T.; Dormann, C.F.; Streif, S.; Jerosch, S.; Götz, M.; et al. Survival and Cause-Specific Mortality of European Wildcat (Felis Silvestris) across Europe. Biol. Conserv. 2021, 261, 109239. [Google Scholar] [CrossRef]
- Pierpaoli, M.; Randi, E.; Ragni, B.; Bir, Z.S.; Szemethy, L.; Herrmann, M.; Hupe, K.; Fernandes, M. Genetic Distinction of Wildcat (Felis Silvestris) Populations in Europe, and Hybridization with Domestic Cats in Hungary. Mol. Ecol. 2003, 12, 2585–2598. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, R.; Godinho, R.; Randi, E.; Ferrand, N.; Alves, P.C. Molecular Analysis of Hybridisation between Wild and Domestic Cats (Felis Silvestris) in Portugal: Implications for Conservation. Conserv. Genet. 2008, 9, 1–11. [Google Scholar] [CrossRef]
- Mattucci, F.; Oliveira, R.; Lyons, L.A.; Alves, P.C.; Randi, E. European Wildcat Populations Are Subdivided into Five Main Biogeographic Groups: Consequences of Pleistocene Climate Changes or Recent Anthropogenic Fragmentation? Ecol. Evol. 2016, 6, 3–22. [Google Scholar] [CrossRef] [Green Version]
- Tiesmeyer, A.; Ramos, L.; Manuel Lucas, J.; Steyer, K.; Alves, P.C.; Astaras, C.; Brix, M.; Cragnolini, M.; Domokos, C.; Hegyeli, Z.; et al. Range-Wide Patterns of Human-Mediated Hybridisation in European Wildcats. Conserv. Genet. 2020, 21, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Cazacu, C.; Adamescu, M.C.; Ionescu, O.; Ionescu, G.; Coţovelea, A.; Jurj, R.; Popa, M.; Cazacu, R. Mapping Trends of Large and Medium Size Carnivores of Conservation Interest in Romania. Ann. For. Res. 2014, 57, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Golemanski, V.; Stoev, P.; Dobrev, D. The Red Data Book of Bulgaria, Volume 2, Animals. Available online: http://e-ecodb.bas.bg/rdb/en/ (accessed on 20 August 2021).
- Halil, I. Red Book of Fauna of the Republic of Kosovo; Ministry of Environment and Spatial Planning Kosovo Institute for Nature Protection: Prishtina, Kosovo, 2019; ISBN 3491446554.
- Legakis, A.; Maragou, P. The Red Book of Endangered Animals of Greece; Legakis, A., Maragou, P., Eds.; Hellenic Zoological Society: Athens, Greece, 2009. [Google Scholar]
- Adamakopoulos, T.; Matsoukas, P. La Distribution Du Chat Sauvage (Felis Silvestris) En Grèce. Biol. Gallo-Hell. 1991, 18, 45–52. [Google Scholar]
- Biró, Z.; Szemethy, L.; Heltai, M. Home Range Sizes of Wildcats (Felis Silvestris) and Feral Domestic Cats (Felis Silvestris f. Catus) in a Hilly Region of Hungary. Mamm. Biol. 2004, 69, 302–310. [Google Scholar] [CrossRef]
- Monterroso, P.; Brito, J.C.; Ferreras, P.; Alves, P.C. Spatial Ecology of the European Wildcat in a Mediterranean Ecosystem: Dealing with Small Radio-Tracking Datasets in Species Conservation. J. Zool. 2009, 279, 27–35. [Google Scholar] [CrossRef]
- Anile, S.; Ragni, B.; Randi, E.; Mattucci, F.; Rovero, F. Wildcat Population Density on the Etna Volcano, Italy: A Comparison of Density Estimation Methods. J. Zool. 2014, 293. [Google Scholar] [CrossRef]
- Harmsen, B.; Foster, R.; Quigley, H. Spatially Explicit Capture Recapture Density Estimates: Robustness, Accuracy and Precision in a Long-Term Study of Jaguars (Panthera Onca). PLoS ONE 2020, 15, e0227468. [Google Scholar] [CrossRef] [PubMed]
- Germain, E.; Benhamou, S.; Poulle, M.L. Spatio-Temporal Sharing between the European Wildcat, the Domestic Cat and Their Hybrids. J. Zool. 2008, 276, 195–203. [Google Scholar] [CrossRef]
- Martín-Díaz, P.; Gil-Sánchez, J.M.; Ballesteros-Duperón, E.; Barea-Azcón, J.M.; Virgós, E.; Pardavila, X.; Moleón, M. Integrating Space and Time in Predator-Prey Studies: The Case of Wildcats and Rabbits in SE Spain. Mamm. Biol. 2018, 88, 114–122. [Google Scholar] [CrossRef]
- Virgós, E.; Tellería, J.L.; Santos, T. A Comparison on the Response to Forest Fragmentation by Medium-Sized Iberian Carnivores in Central Spain. Biodivers. Conserv. 2002, 11, 1063–1079. [Google Scholar] [CrossRef]
- Lozano, J. Habitat Use by European Wildcats (Felis Silvestris) in Central Spain: What Is the Relative Importance of Forest Variables? Anim. Biodivers. Conserv. 2010, 33, 143–150. [Google Scholar]
- Mangas, J.G.; Lozano, J.; Cabezas-Díaz, S.; Virgós, E. The Priority Value of Scrubland Habitats for Carnivore Conservation in Mediterranean Ecosystems. Biodivers. Conserv. 2008, 17, 43–51. [Google Scholar] [CrossRef]
- Jerosch, S.; Kramer-Schadt, S.; Götz, M.; Roth, M. The Importance of Small-Scale Structures in an Agriculturally Dominated Landscape for the European Wildcat (Felis Silvestris Silvestris) in Central Europe and Implications for Its Conservation. J. Nat. Conserv. 2018, 41, 88–96. [Google Scholar] [CrossRef]
- Chatziantoniou, A.; Petropoulos, G.P.; Psomiadis, E. Co-Orbital Sentinel 1 and 2 for LULC Mapping with Emphasis on Wetlands in a Mediterranean Setting Based on Machine Learning. Remote Sens. 2017, 9, 1259. [Google Scholar] [CrossRef] [Green Version]
- Potoçnik, H.; Kljun, F.; Raçnik, J.; Skrbinšek, T.; Adamiç, M.; Kos, I. Experience Obtained from Box Trapping and Handling Wildcats in Slovenia. Acta Theriol. 2002, 47, 211–219. [Google Scholar] [CrossRef]
- Anile, S.; Bizzarri, L.; Lacrimini, M.; Sforzi, A.; Ragni, B.; Devillard, S. Home-Range Size of the European Wildcat (Felis Silvestris Silvestris): A Report from Two Areas in Central Italy. Mammalia 2017, 82, 1–11. [Google Scholar] [CrossRef]
- Jerosch, S.; Götz, M.; Roth, M. Spatial Organisation of European Wildcats (Felis Silvestris Silvestris) in an Agriculturally Dominated Landscape in Central Europe. Mamm. Biol. 2017, 82, 8–16. [Google Scholar] [CrossRef]
- Kitchener, A.C.; Yamaguchi, N.; Ward, J.M.; Macdonald, D.W. A Diagnosis for the Scottish Wildcat (Felis Silvestris): A Tool for Conservation Action for a Critically-Endangered Felid. Anim. Conserv. 2005, 8, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Kilshaw, K.; Drake, A.; Macdonald, D.W.; Kitchener, A.C. The Scottish Wildcat: A Comparison of Genetic and Pelage Characteristics; Scottish Natural Heritage Commisioned Report No. 356; Scottish Natural Heritage: Scotland, UK, 2010; 70p. [Google Scholar]
- Kenward, R. A Manual for Wildlife Radio Tagging, 2nd ed.; Academic Press: New York, NY, USA, 2001. [Google Scholar]
- Horne, J.S.; Garton, E.O.; Krone, S.M.; Lewis, J.S. Analyzing Animal Movements Using Brownian Bridges. Ecology 2007, 88, 2354–2363. [Google Scholar] [CrossRef]
- Team, R.C. A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 22 August 2021).
- Nielson, R.M.; Sawyer, H.; McDonald, T.L. BBMM: Brownian Bridge Movement Model. R Package Version 3.0. Available online: https://cran.r-project.org/package=BBMM (accessed on 25 August 2021).
- Calenge, C. The Package “Adehabitat” for the R Software: A Tool for the Analysis of Space and Habitat Use by Animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Wikelski, M.; Davidson, S.; Kays, R. Movebank: Archive, Analysis and Sharing of Animal Movement Data. Hosted by the Max Planck Institute of Animal Behavior. Available online: www.movebank.org (accessed on 10 July 2021).
- Wilson, R.P.; White, C.R.; Quintana, F.; Halsey, L.G.; Liebsch, N.; Martin, G.R.; Butler, P.J. Moving towards Acceleration for Estimates of Activity-Specific Metabolic Rate in Free-Living Animals: The Case of the Cormorant. J. Anim. Ecol. 2006, 75, 1081–1090. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference—A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Barton, K.; Barton, M.K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. Version 2020, 1, 18. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. LmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Byrnes, J. Re: Coefficient of Determination (R^2) When Using Lme(). Available online: https://stat.ethz.ch/pipermail/r-sig-mixed-models/2008q2/000713.html (accessed on 23 August 2021).
- QGIS.org. QGIS Geographic Information System, QGIS Association. Available online: http://www.qgis.org (accessed on 22 August 2021).
- European Union’s Earth Observation Program “Copernicus”. Available online: www.copernicus.eu (accessed on 26 August 2021).
- Burgman, M.A.; Fox, J.C. Bias in Species Range Estimates from Minimum Convex Polygons: Implications for Conservation and Options for Improved Planning. Anim. Conserv. 2003, 6, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Bizzarri, L.; Lacrimini, M.; Ragni, B. Live Capture and Handling of the European Wildcat in Central Italy. Hystrix 2010, 21. [Google Scholar] [CrossRef]
- Dötterer, M.; Bernhart, F. The Occurance of Wildcats in the Southern Swiss Jura Mountains. Acta Theriol. 1996, 41, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Daniels, M.J.; Beaumont, M.A.; Johnson, P.J.; Balharry, D.; Macdonald, D.W.; Barratt, E. Ecology and Genetics of Wild-Living Cats in the North-East of Scotland and the Implications for the Conservation of the Wildcat. J. Appl. Ecol. 2001, 38, 146–161. [Google Scholar] [CrossRef]
- Oliveira, T.; Urra, F.; López-Martín, J.M.; Ballesteros-Duperón, E.; Barea-Azcón, J.M.; Moléon, M.; Gil-Sánchez, J.M.; Alves, P.C.; Díaz-Ruíz, F.; Ferreras, P.; et al. Females Know Better: Sex-Biased Habitat Selection by the European Wildcat. Ecol. Evol. 2018, 8, 9464–9477. [Google Scholar] [CrossRef]
- Ruiz-Olmo, J.; Pinyol, C.; Sánchez, D.; Such-Sanz, À. Breeding Pattern of Wildcat Felis Silvestris (Schreber, 1777) Studied in Captivity in the Iberian Peninsula. Hystrix 2018, 29, 202–210. [Google Scholar] [CrossRef]
- Sarmento, P.; Cruz, J.; Tarroso, P.; Fonseca, C. Space and Habitat Selection by Female European Wild Cats (Felis Silvestris Silvestris). Wildl. Biol. Pract. 2006, 2, 79–89. [Google Scholar] [CrossRef]
- Corbett, L.K. Feeding Ecology and Social Organization of Wildcats (Felis Silvestris) and Domestic Cats (Felis Catus) in Scotland. Ph.D. Thesis, University of Aberdeen, Aberdeen, Scotland, UK, 1979. [Google Scholar]
- Kyba, C.C.M.; Mohar, A.; Posch, T. How Bright Is Moonlight? Astron. Geophys. 2017, 58, 1.31–1.32. [Google Scholar] [CrossRef]
- Botts, R.T.; Eppert, A.A.; Wiegman, T.J.; Blankenship, S.R.; Rodriguez, A.; Wagner, A.P.; Ullrich, S.E.; Allen, G.R.; Garley, W.M.; Asselin, E.M.; et al. Does Moonlight Increase Predation Risk for Elusive Mammals in Costa Rica? Trop. Conserv. Sci. 2020, 13. [Google Scholar] [CrossRef]
- Michalski, F.; Norris, D. Activity Pattern of Cuniculus Paca (Rodentia: Cuniculidae) in Relation to Lunar Illumination and Other Abiotic Variables in the Southern Brazilian Amazon. Zoologia 2011, 28, 701–708. [Google Scholar] [CrossRef] [Green Version]
- Pratas-Santiago, L.P.; Gonçalves, A.L.S.; Nogueira, A.J.A.; Spironello, W.R. Dodging the Moon: The Moon Effect on Activity Allocation of Prey in the Presence of Predators. Ethology 2017, 123, 467–474. [Google Scholar] [CrossRef]
- Huck, M.; Juárez, C.P.; Fernández-Duque, E. Relationship between Moonlight and Nightly Activity Patterns of the Ocelot (Leopardus Pardalis) and Some of Its Prey Species in Formosa, Northern Argentina. Mamm. Biol. 2017, 82, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Jerosch, S.; Götz, M.; Klar, N.; Roth, M. Characteristics of Diurnal Resting Sites of the Endangered European Wildcat (Felis Silvestris Silvestris): Implications for Its Conservation. J. Nat. Conserv. 2010, 18, 45–54. [Google Scholar] [CrossRef]
- Tatler, J.; Cassey, P.; Prowse, T.A.A. High Accuracy at Low Frequency: Detailed Behavioural Classification from Accelerometer Data. J. Exp. Biol. 2018, 221, jeb184085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kröschel, M.; Reineking, B.; Werwie, F.; Wildi, F.; Storch, I. Remote Monitoring of Vigilance Behavior in Large Herbivores Using Acceleration Data. Anim. Biotelem. 2017, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Rast, W.; Kimmig, S.E.; Giese, L.; Bergerid, A. Machine learning goes wild: Using data from captive individuals to infer wildlife behaviours. PLoS ONE 2020, 15, e0227317. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Sex | Age (Years) | Weight (kg) | Capture Date | Last Relocation Date | Days Tracked | GPS Fixes | 2-min Acc/Meter Logs |
|---|---|---|---|---|---|---|---|---|
| Apollo | M | 2–3 | 2.9 | 29 September 2020 | 5 January 2021 | 99 | 3976 | 70,190 |
| Aphrodite | F | 2–3 | 3.6 | 1 October 2020 | 18 June 2021 | 261 | 11,951 | 170,893 |
| Ares | M | 2–3 | 2.6 | 2 October 2020 | 1 June 2021 | 243 | 11,845 | 167,349 |
| Artemis | F | 1–2 | 2.7 | 30 October 2020 | 23 June 2021 | 237 | 11,278 | - |
| Ira | F | 3–4 | 3.7 | 4 November 2020 | 30 June 2021 | 239 | 11,875 | 171,679 |
| Aphrodite | Artemis | Ira | Apollo | Ares | Mean | ||
|---|---|---|---|---|---|---|---|
| TOTAL | BB | 0.94 | 3.08 | 0.97 | 1.22 | 4.43 | 2.13 ± 1.40 |
| MCP | 1.69 | 4.79 | 1.77 | 2.35 | 58.37 | 13.79 ± 22.32 | |
| October–December | BB | 1.17 | 1.10 | 1.13 | 1.54 | 1.64 | 1.32 ± 0.23 |
| MCP | 2.10 | 1.12 | 1.43 | 2.07 | 3.73 | 2.09 ± 0.90 | |
| January–March | BB | 0.50 | 2.01 | 0.87 | 0.51 1 | 3.08 | 1.62 ± 1.012 |
| MCP | 0.44 | 5.01 | 0.96 | 0.83 1 | 37.87 | 11.07 ± 15.572 | |
| April–June | BB | 0.84 | 1.90 | 0.67 | - | 2.07 | 1.37 ± 0.622 |
| MCP | 1.20 | 3.31 | 0.88 | - | 8.15 | 3.39 ± 2.912 |
| Variables | Estimate | SE | z-Value | Pr(>|z|) |
|---|---|---|---|---|
| intercept (βο) | 3963.3 | 207.0 | 19.122 | <0.0001 |
| day length | −2086.8 | 321.4 | 6.484 | <0.0001 |
| moon phase | 89.6 | 58.8 | 1.522 | 0.128 |
| min. temperature | 20.2 | 4.5 | 4.463 | <0.0001 |
| Variables | Estimate | SE | df | t-Value | Pr(>|z|) |
|---|---|---|---|---|---|
| intercept (βο) | 4582.1 | 395.1 | 22.918 | 11.598 | <0.0001 |
| day length | −2814.1 | 690.9 | 797.93 | −4.073 | <0.0001 |
| min. temperature | 81.2 | 9.7 | 796.819 | 8.333 | <0.0001 |
| Individual | Sex | Unique Sites | October | November | December | January | February | March | April | May | June |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Aphrodite | F | 23 | 5 | 4 | 4 | 4 | 6 | 4 | 5 | 2 | - |
| Ira | F | 19 | - | 5 | 3 | 6 | 3 | 5 | 4 | 1 | 3 |
| Apollo | M | 9 | 5 | 1 | 3 | - | - | - | - | - | - |
| Ares | M | 23 | 4 | 4 | 4 | 4 | 3 | 5 | 5 | 3 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migli, D.; Astaras, C.; Boutsis, G.; Diakou, A.; Karantanis, N.-E.; Youlatos, D. Spatial Ecology and Diel Activity of European Wildcat (Felis silvestris) in a Protected Lowland Area in Northern Greece. Animals 2021, 11, 3030. https://doi.org/10.3390/ani11113030
Migli D, Astaras C, Boutsis G, Diakou A, Karantanis N-E, Youlatos D. Spatial Ecology and Diel Activity of European Wildcat (Felis silvestris) in a Protected Lowland Area in Northern Greece. Animals. 2021; 11(11):3030. https://doi.org/10.3390/ani11113030
Chicago/Turabian StyleMigli, Despina, Christos Astaras, George Boutsis, Anastasia Diakou, Nikolaos-Evangelos Karantanis, and Dionisios Youlatos. 2021. "Spatial Ecology and Diel Activity of European Wildcat (Felis silvestris) in a Protected Lowland Area in Northern Greece" Animals 11, no. 11: 3030. https://doi.org/10.3390/ani11113030